
Abstract
Ectotherms display substantial demographic variation across latitudinal gradients of temperature. Higher temperatures are often associated with smaller size, rapid initial growth rates, and early maturation, generally described as the Temperature-Size Rule (TSR). The longevity of most ectotherms also declines at warmer, lower latitudes. However, these patterns may be modified by increases in food resources that can flow on to continuous growth and large adult size. The present study estimates age-based demographic parameters of large-bodied tropical wrasses (Hemigymnus melapterus, H. fasciatus, Cheilinus fasciatus, and Oxycheilinus digramma) collected from Philippine fish markets (9–11°N) and sampled from Palm (18.53–18.70°S) and Whitsunday (20.05–20.21°S) reefs on the Great Barrier Reef, Australia (GBR). Differences in longevity, initial growth rates, and the age at sexual maturation at a biogeographic scale, between the Philippines and GBR, conformed to predictions of the TSR. However, Philippine specimens exhibited greater relative body condition and sustained periods of growth beyond sexual maturity resulting in larger adult size than GBR samples. Size-structure data from Philippine marine reserves and fished sites indicated that these differences were not confounded by fishery-dependent sampling. Moreover, latitudinal length–weight relationships could not be explained by lower densities of the focal wrasses in the Philippines or by relative gonad size. Less pronounced patterns of demographic variation that differed across species were evident at a regional scale, among Palm and Whitsunday reefs. Patterns of demographic variation between the Philippines and GBR strongly suggest that differences in food resource levels will be important in explaining the observed geographic variation.
Similar content being viewed by others
Introduction
Life‐history variation in ectotherms arises from trade‐offs that enhance reproductive outputs at the cost of somatic maintenance and survival. Age and body size at maturity are particularly important life history traits, as they affect most biological rates and are principal determinants of individual fitness (Stearns 1992; Perez and Munch 2010). These traits and the nature and magnitude of their variation are thus critical elements in the explanation of demographic trade-offs that result from the response of individuals and their offspring to environmental variation.
Ectotherms generally display substantial life‐history clines across latitudinal gradients of temperature. Ectotherms at high latitudes often attain a greater longevity relative to conspecifics at warmer, lower latitudes, and this is thought to result from the associated effects of temperature on metabolism (consistent with the Metabolic Theory of Ecology; Brown et al. 2004; Munch and Salinas 2009). Laboratory experiments have also associated lower relative temperatures with slower initial growth rates and larger adult size across a broad range of ectotherms (from bacteria to vertebrates), often referred to as the Temperature-Size Rule (TSR: Atkinson 1994). Nevertheless, in real-world systems, temperature co-varies with numerous other factors known to influence life histories, such as resource levels, and these environmental co-variates can modify the temperature-related growth patterns of ectotherms (Huey and Kingsolver 2019). This is particularly true across coral reef environments which are (i) biologically variable at a range of spatial scales; (ii) subject to frequent disturbances that may profoundly modify habitats; and (iii) populated by species that have very broad geographic distributions and complex life histories, including protogynous hermaphroditism. For these reasons, the demographic changes seen in coral reef fishes distributed across latitudinal gradients may not align with predictions based on laboratory experiments, but rather may reflect interactions between temperature and other environmental co-variates (Munch and Salinas 2009; Audzijonyte et al. 2019; Lowe et al. 2021a). By identifying major demographic departures from the predictions of the TSR across latitudinal scales and postulating the factor responsible based on the known effects of other co-variates on the life histories of ectotherms, studies can inform mechanistic hypotheses aimed at determining whether the organism is responding to the factor of interest, the indirect effect of a co-variate, or the interactive effects of several factors.
The primary research goals are therefore (1) to document how age-based life history features in coral reef fishes vary over latitudinal gradients, (2) to assess whether patterns of life history variation reflect the characteristic reaction norms of most ectotherms to variation in temperature, (3) to identify areas of demographic departure from expected effects of temperature and suggest the underlying processes based on known effects of other co-variates on life histories, and (4) suggest hypotheses which mechanistically test the importance of the factor thought responsible for the demographic departure from other co-variates acting on life histories. Furthermore, as most management decisions concerning harvested fish populations seek to incorporate rate processes (e.g., growth, recruitment, mortality etc.), an important element is the collection of age-based information which allows estimates of demographic rates.
The present study documents the variability in age-based demographic characteristics of four large-bodied tropical wrasses (Hemigymnus melapterus, H. fasciatus, Cheilinus fasciatus, and Oxycheilinus digramma) at a biogeographic scale spanning 30° of latitude, between the Philippines and the Great Barrier Reef, Australia (GBR), and at a regional scale, between Palm and Whitsunday reefs of the GBR separated by 2° of latitude. We then assess whether patterns of life history variation correlate with the predictions of the TSR and postulate the factors responsible for any demographic departures to inform mechanistic hypotheses and future sampling efforts. These species are large-bodied (maximum length 30–60 cm) and abundant, mobile predators of benthic invertebrates on coral reefs in the Indo-Pacific (Kramer et al. 2015). In developing countries, such as the Philippines, these wrasses are commonly captured by spear, trap, hook and line, and gill nets in small-scale fisheries (Russ and Alcala 1998). Conversely, in Australia, the four study species are not targeted by recreational or commercial fisheries. Demographic consequences of environmental change on the focal wrasses have yet to be quantified. However, it is notable that the sexual ontogenies of these species differ among Philippine and GBR reefs, with H. melapterus, C. fasciatus, and O. digramma exhibiting evidence of functional gonochorism and diandric protogyny on Philippine reefs, while on GBR reefs these three species plus H. fasciatus showed evidence of monandric protogynous hermaphroditism (Lowe et al. 2021b).
Fishery-dependent and fishery-independent sampling was used to determine life history traits (growth rates, mortality rates, body condition, long-term density, mean and maximum size and age, age and size at maturity and sex change) in the four focal wrasse species. Wrasses were collected from fish markets in the central Philippines, and from populations at two unfished locations on the Great Barrier Reef (GBR), Australia. By describing and discussing patterns of wrasse life history trait variation across biogeographic (30°) and regional (2°) latitudinal distances, we provide important demographic information which will contribute toward future causative studies of the drivers of life history variation in these species across various spatial scales. Based on studies of other labrids and the perceived metabolic costs of living at low latitudes, we expect that wrasse growth rates will increase with increasing temperature, while longevity, maximum and asymptotic body length, and the age and length of maturation will decline on low latitude reefs (Atkinson 1994; Munch and Salinas 2009; Lek et al. 2012; Trip et al. 2014).
Methods
Sampling locations
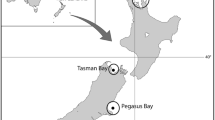
Hemigymnus melapterus, H. fasciatus, C. fasciatus, and O. digramma were collected from fringing coral reefs surrounding the Palm and Whitsunday Island groups in the central GBR region, Australia and purchased from fish markets in the Philippines (Fig. 1). GBR samples were collected between 2016 and 2019 using spearguns on scuba and individual fish were collected without preference for body size. Collection sites were positioned on fringing reefs surrounding Pelorus and Curacoa in the Palm Island group, and Hook and Whitsunday in the Whitsunday Island group (Fig. 1b, c). Philippine samples were purchased primarily from fish markets in Dumaguete, Negros Island (Fig. 1a). Adult wrasse samples from the Philippines were often caught using spearguns or in fish traps, while juveniles were commonly caught using gill nets. It was not possible to track the source reef of each purchased wrasse specimen in the Philippines, but conversations with fish vendors indicated supply chains that were quite short (reefs in the vicinity of Negros) as well as specimens that originated from reefs as far as 300–500 km away (e.g., Palawan) (Fig. 1a).

Maps indicating primary collection sites and likely catchment area of fishery-dependent samples collected in the Philippines (A), and areas of fishery-independent sampling at the Palm (B) and Whitsunday (C) Island Groups, GBR, Australia
Age determination
In total, 309 H. melapterus, 227 C. fasciatus, and 216 O. digramma were collected from the Great Barrier Reef, Australia and from fish markets in the Philippines (see Table 1 for location-specific sample sizes). Despite being common on Philippine reefs, Hemigymnus fasciatus was rare in fish markets and an insufficient number of samples were collected for the estimation of life history parameters for this species in the Philippines. Consequently, all life history parameters for H. fasciatus were estimated from 115 fish collected on GBR reefs. Once samples were collected, fish were placed on ice and processed immediately on return to the processing area. The body size of both Hemigymnus spp. and O. digramma were recorded as total length (TL; nearest mm) due to their rounded caudal fin. Conversely, fork length (FL; nearest mm) was used for C. fasciatus due to its emarginate caudal fin with pronounced dorsal and ventral lobes (in adults). Total weight (W; nearest 0.01 g in Australia, 1 g in Philippines) was recorded for each specimen. Otoliths were removed, cleaned, and stored dry at the time of processing. Subsequently, one otolith from each sagittal pair was weighed to the nearest 0.0001 g on the GBR and 0.001 g in the Philippines for analysis of otolith weight and age (ESM 1). Age was estimated for each individual through interpretation of putative annual and daily increments in the microstructure of sectioned sagittal otoliths (Fig. 2), see Lowe et al. (2021b) for details.

Transverse sections of sagittal otoliths displaying putative annual increments: a Hemigymnus melapterus, b H. fasciatus, c Cheilinus fasciatus, d Oxycheilinus digramma. White lines indicate annual bands. Otolith sections are from GBR populations, images are not to one scale
Statistical analysis
To estimate life history parameters from length-at-age data, separate von-Bertalanffy growth functions (VBGFs) were fit for each species at each location using the FSA package in R (Ogle et al. 2019). The original parametrization of the VBGF,\(Lt = L\infty - \left( {L\infty - L_0 } \right)e^{ - Kt}\), was used to determine estimates of K and L∞ at a common length at settlement (L0). Lt is the length (TL or FL) of a fish at age t (years), L∞ is the mean asymptotic body length, K describes the curvature of growth toward L∞, and L0 is the length at which t = 0. Settlement length (L0) was fixed at 10 mm to improve parameter estimates and was based on the approximate larval duration of the focal wrasses multiplied by an average pre-settlement growth rate determined for Indo-Pacific wrasses (Victor 1986). Estimates of t0 (the theoretical age at which Lt = 0) were extrapolated using non-linear least squares regression from the original VBGF fit with a fixed L0. Length-at-age data were bootstrapped to produce 1000 samples of parameter estimates for each species, at each location.
To quantify differences in growth rates between Palm, Whitsunday, and Philippine wrasse populations, the Francis VBGF parametrization (rVBGF) was also fit to size-at-age data using the FSA package in R (Ogle et al. 2019). The rVBGF differs from the VBGF in that ages (τ, ω, and μ) and their corresponding mean lengths (Lτ, Lω, and Lμ) can be specified within the model. The rVBGF is represented by \(Lt = L_\tau + \left( {L\mu - L\tau } \right) \frac{{1 - r^2 \frac{{\left( {t - \tau } \right)}}{{\left( {\mu - \tau } \right)}}}}{1 - r^2 }\), where \(r = \frac{L\mu - L\omega }{{L\omega - L\tau }}\). For H. melapterus, H. fasciatus, and C. fasciatus, ages τ, ω, and μ were set at 1, 3, and 5 years, respectively, to estimate initial, mid-life, and approximate asymptotic growth of these species across locations. For O. digramma, τ, ω, and μ were set at 1, 2, and 3 years, respectively, as the growth of Palm populations was largely asymptotic by the age of 3, and only one individual was recorded > 3 yrs in the Philippines. 1000 bootstrap estimates of Lτ, Lω, and Lμ parameters were computed for each species at each location. Mean maximum age (Tmax) and mean maximum length (Lmax) of the oldest and largest 15% of the sample at each location were also calculated.
Estimates of instantaneous total mortality (Z) were fitted with the FSA package in R as the absolute value of the slope from a linear regression fit to the natural logarithm of age frequency (Ogle et al. 2019). Fish were considered fully recruited to the sampling regime at the modal age of samples obtained in each location and age classes without samples were excluded from the analysis. Regressions used to estimate mortality were compared between locations via analysis of covariance (ANCOVA) and subsequent Tukey’s Honestly Significant Difference (HSD) tests.
Designations of sex and stage from histological examination of gonads followed Lowe et al. (2021b). The timing of female maturity was estimated by fitting a logistic curve to the proportional frequency of immature to mature females in 2 cm size class bins and 1-year age class bins. The same protocol was used to estimate the age and size of sex change using the proportional frequency of females to males per size or age bin. Reproductive life history traits estimated via this method were the length of 50% and 90% female maturity (Lm50, Lm90), age of 50% and 90% female maturity (Tm50, Tm90), length of 50% and 90% sex change (Ls50, Ls90), and the age of 50% and 90% sex change (Ts50, Ts90).
The relative timing of maturity and sex change were examined by dividing Lm50, Tm50, Ls50, and Ts50 estimates by the respective mean maximum length (Lmax) and age (Tmax) of each species in each location.
Separate body length–weight and otolith weight to age relationships were estimated for each species at each location. Length–weight relationships were routinely estimated in the form w = αLb via a logarithmic transformation of each individual’s length (L) and weight (w), with α (intercept) and b (slope) estimated by ordinary least squares regression. Length–weight relationships were compared for significant differences using ANCOVA’s and Tukey HSD tests. To determine relative differences in fish condition with size between locations, a generalized logarithmic length–weight model was fit to all populations. Residuals for each individual were then extracted, plotted with body size, and fit with linear models. Species-specific otolith weight to age relationships at each location were modeled using linear regressions.
Long-term species-specific densities were compared between GBR and Philippine reefs to determine whether lower densities correlated with greater growth rates, body condition, maximum or asymptotic size, as would be expected under density dependent competition for food resources. Wrasse densities in the Philippines were surveyed 26 times in 31 years, between 1983 and 2014, by a single observer (GRR) using six replicate 50 m × 20 m underwater visual census (UVCs) conducted on the reef slopes (9–17 m) at two sites, Apo and Sumilon Island fished areas, which are located in the immediate catchment area of markets sampled in south-eastern Negros. Wrasse densities in Australia were surveyed biennially for 12 years, from 2007 to 2018, by conducting five replicate 50 m × 6 m UVC transects on reef slopes (4–12 m in depth) at each of the 71 survey sites positioned adjacent to collection sites at the Palm (30 survey sites) and Whitsunday (41 survey sites) Island Groups. Long-term wrasse densities were calculated for each location as the mean species-specific density observed across pooled Philippine and GBR sites for the entire survey period of 31 years and 12 years, respectively. Full details of survey methodology can be found in Russ et al. (2017) and Lowe et al. (2019).
To assess whether fishery-dependent sampling biased the length-based life history estimates of Philippine wrasses in the present study, species-specific size structure observed during visual surveys conducted across 19 no-take marine reserves and 31 fished sites in the Bohol Sea (i.e., the immediate catchment area of the fish markets sampled in Negros Island) were pooled and compared. Surveys were conducted on scuba from 2006 to 2008 on the reef crests and slopes of several large and small islands (Bohol, Negros, southern Leyte, Siquijor, North-west Mindanao, Selinog and Aliguay) and in 2019 at Apo and Sumilon Islands. All sites were surveyed using 50 m long × 10 m wide transects, except Apo and Sumilon where 50 m long × 20 m replicates were used (n = 312 transects/replicates; reserves = 120, fished = 192). Surveys at each site were undertaken by a single fish observer who counted and estimated the TL of fish to the nearest 1 cm. Estimation of fish size to the nearest 1 cm was possible in Apo and Sumilon because the surveys were conducted along three 50-m long strips that were about 7 m wide (see Russ and Alcala 1996 for full methods). Size estimates were later compiled into 3 cm size categories and plotted to inspect trends. Summary statistics calculated for size structure data included maximum size, which was determined as the largest individual observed, mean maximum size, which was calculated identically to Lmax in the present study (i.e., the mean size of the largest 15% of individuals observed), and the mean size of all individuals observed. Mean sizes observed in reserves and fished sites were compared via ANOVA.
Results
Growth increments were evident in H. melapterus, H. fasciatus, C. fasciatus, and O. digramma otoliths from GBR and Philippine populations (Fig. 2). Optical clarity and interpretability increased from low to high latitudes, and across all locations only 2.43% of samples were deemed unreadable and discarded from analysis. Otolith weight to fish age relationships indicated that Philippine populations of H. melapterus, C. fasciatus, and O. digramma often had heavier otoliths per age class compared to samples obtained from GBR reefs (ESM 1). Otolith increment counts were in relatively good agreement, with 70.15% of the first two counts being identical, 25.02% of reads differed by 1 year, and 3.88% differed by 2–4 years.
Geographic variation in growth, maturity, and sex change
Table 1 summarizes the sample size, mortality rate, size range, growth, and longevity estimates for each species at each location. Sizes at maturity and sex change for each species at each location are presented in Table 2. Variations in life histories were evident at both biogeographic (between Philippine and GBR populations) and regional scales (between inshore GBR locations). Three trends were clear across species: (1) longevity was greater on GBR reefs than on Philippine reefs (Table 1; Fig. 3); (2) Philippine wrasses grew faster, matured earlier, and experienced a higher mortality rate than those on unfished GBR reefs (Tables 1, 2; Figs. 3, 4, 5, 6); (3) despite this, Philippine wrasses were heavier for a given size (i.e., had greater relative body condition) and achieved a similar asymptotic and maximum length to populations on the GBR (Table 1; Figs. 3, 6, 7).

von-Bertalanffy growth function (VBGF) curves fit to size-at-age data for a H. melapterus, b H. fasciatus, c C. fasciatus, and d O. digramma specimens collected from the Whitsunday and Palm Island groups, GBR, and sampled from Philippine fish markets. Data points and colors appear as per legend. Parameter estimates are in Table 1, individual growth curves and size-at-age data for each species in each location are in ESM 1

Relative timing of maturity (a, b) and sex change (c, d) of H. melapterus, H. fasciatus, C. fasciatus, and O. digramma at the Whitsunday and Palm Island groups, GBR, and from Philippine fish markets. Data points and colors appear as per legend

Bootstrapped 95% confidence ellipses surrounding rVBGF estimates of Lτ and Lμ, describing growth from year 1 until approximate asymptotic size for each species at each location. Data points and colors appear as per legend

Age-based catch curves for a H. melapterus, b H. fasciatus, c C. fasciatus, and d O. digramma on Palm and Whitsunday reefs (left column), Philippine reefs (right column). A modal age of 1 was used for mortality calculations of Whitsunday H. fasciatus populations, despite anomalously high age class frequencies of ages 7 and 8 (explained in Results). Gray data points were excluded from catch curve analysis. 95% confidence intervals for mortality estimates appear in Table 1

Length–weight relationships of a H. melapterus, b H. fasciatus, c C. fasciatus, d O. digramma, collected from Palm and Whitsunday Island groups, GBR and from Philippine fish markets, e linear regressions comparing relative condition at body length across locations and species, f length–weight model parameters for each species at each location. * indicates a statistically significant difference between GBR and Philippine reefs
When differences in the mean maximum length (Lmax) and mean maximum age (Tmax) of populations were accounted for, 95% confidence intervals surrounding the relative size and age of maturity overlapped across species sampled from GBR reefs (Fig. 4a, b), all of which are monandric protogynous hermaphrodites. Similarly, the 95% confidence intervals surrounding the relative size and age of maturity overlapped across species sampled from Philippine reefs (Fig. 4a, b), whereby all species exhibited histological evidence of functional gonochorism and diandric protogyny. Estimates of the relative size and age of maturity did not consistently overlap between Philippine and GBR reefs (Fig. 4a, b), albeit this was expected given the different patterns of sexual ontogeny in these localities. These data indicate that, depending on the location, most of these wrasse populations reach sexual maturity (Lm50) at 47% – 62% of their respective mean maximum length, with H. melapterus and C. fasciatus in the Philippines lying well outside this, reaching sexual maturity at 21% and 32% of Lmax, respectively (Fig. 4a). The 95% confidence intervals surrounding estimates of the relative age and size of sex change also overlapped for H. melapterus, H. fasciatus, and C. fasciatus sampled from Palm and Whitsunday reefs of the GBR, and estimates of the relative size of sex change were very similar between GBR reefs for O. digramma, albeit 95% confidence intervals did not overlap (Fig. 4c, d). Conversely, the relative size and age at sex change among Philippine taxa were far from invariant (Fig. 4c, d), despite these species undergoing similar patterns of sexual ontogeny on Philippine reefs. Overall, most of the wrasse populations achieved sex change (Ls50) between 77% and 89% of each population’s respective mean maximum length, with the exception of Philippine C. fasciatus and O. digramma, which were found to change sex at 32% and 53% of population-specific Lmax, respectively (Fig. 4c).
Mortality
Mortality rates on GBR reefs were similar for all species investigated (ANCOVA, p > 0.05) and fished Philippine wrasse populations had higher mortality rates than unfished GBR populations (Table 1; Fig. 6). Differences between Whitsunday and Philippine mortality rates were significant for C. fasciatus (ANCOVA, p = 0.015; Tukey HSD, p = 0.002), whereas Palm and Philippine mortality rates were not significantly different for this species (Tukey HSD, p = 0.06). Hemigymnus melapterus and O. digramma had higher mortality rates in the Philippines (Fig. 6), but differences between mortality slopes were not statistically significant (ANCOVA, p = 0.076 and 0.21, respectively). Anomalously high age class frequencies of age 7 and 8 H. fasciatus were sampled from the Whitsunday Island group, likely due to a strong year of recruitment in 2007/2008 (Lowe et al. 2020). However, a modal age of 1 was used in mortality estimates, rather than age 8, as fitting a regression from ages 8 onwards would result in an unrealistic mortality rate for an unfished population of H. fasciatus. This was particularly so, considering mortality rate estimates for the same species at the Palm Island group, and the relative difference between Palm and Whitsunday mortality rates for other species studied here (Fig. 6; Table 1).
Length–weight and body condition relationships
Length–weight relationships of all four species indicated allometric growth (b ≠ 3) in all locations (Fig. 7). Philippine H. melapterus and C. fasciatus individuals were significantly heavier for a given length than those on Palm and Whitsunday reefs, GBR (Fig. 7a, c; ANCOVA, p < 0.05; Tukey HSD, p < 1*e−4, and < 1*e−5, respectively). This was particularly clear for large-bodied fish (Fig. 7a, c, e). Philippine O. digramma were significantly heavier for a given length than those on Whitsunday reefs, GBR (ANCOVA, p < 0.05; Tukey HSD, p < 1*e−4) but were not significantly different to those on Palm reefs (Tukey HSD; p = 0.093). Hemigymnus melapterus, H. fasciatus, C. fasciatus, and O. digramma did not exhibit a significant difference in length–weight relationships between Palm and Whitsunday reefs, GBR (Fig. 7a–d; Tukey HSD, p > 0.05). Linear models fit to relative estimates of body condition (described in “Methods”) indicated that significant differences in body condition begin to occur at approximately 150 mm length across species on Philippine and Australian reefs, whereby wrasses on Philippine reefs were significantly heavier than those on the GBR for a given size (Fig. 7e; Tukey HSD, p = < 1e−5).
Long-term density
While H. melapterus and C. fasciatus were significantly more abundant on GBR reefs compared to those of the Philippines (Fig. 8; Wilcoxon rank-sum test, p = 0.0003 and p = 3.595e−8, respectively), long-term densities of O. digramma were significantly greater in the Philippines than on the GBR (Fig. 8; Wilcoxon rank-sum test, p = 0.0005). As such, wrasse densities were not consistently lower on fished Philippine reefs compared to unfished GBR reefs (Fig. 8).

Raincloud plot showing long-term densities of H. melapterus, C. fasciatus, and O. digramma, calculated over 31 years from Apo and Sumilon Island fished areas, Philippines (red) and over 12 years at the Palm and Whitsunday Island Groups, GBR, Australia (blue). Individual points show mean density per sampled year, boxplots extend from minimum to maximum values, and the probability distribution is shown by shaded bands. Horizontal lines in boxplots indicate the long-term median density of each species in each location. A * next to a species name indicates a significant difference between locations
Size-structure estimates from Philippine Marine reserves and fished control sites
Species-specific size structure estimates pooled across 19 no-take marine reserves and 31 fished sites in the immediate catchment area of fish markets sampled in Negros Island, Central Philippines, revealed that H. melapterus and C. fasciatus achieved larger maximum sizes in fished areas than in marine reserves, while O. digramma had a slightly smaller maximum size in reserves (Fig. 9a–d). Nevertheless, the former two wrasse species had identical estimates of mean maximum size in reserve and fished areas, while mean maximum size of O. digramma was 20.8 cm in reserves and 20 cm in fished sites (Fig. 9d). All species achieved a larger relative mean size inside reserves (Fig. 9d), and this difference was statistically significant for H. melapterus and O. digramma but not C. fasciatus (ANOVA, p = 0.0119, p = 0.032, and p = 0.2, respectively).

Pooled size structure of a Hemigymnus melapterus, b Cheilinus fasciatus, and c Oxycheilinus digramma observed via visual census conducted across 19 marine reserves and 31 fished sites in the Bohol Sea (immediate catchment area of fish markets sampled in Negros Island) during 2006–2008 and 2019. Key summary statistics for each species are shown in d. Colors appear as per legend in a
Estimates of maximum size and mean maximum size from visual surveys of marine reserves and fished sites were very similar to estimates of Lmax and maximum size estimated during the present study (Fig. 9d; Table 1). The mean maximum sizes of H. melapterus and O. digramma observed during visual surveys were within the 95% CIs of Lmax estimates provided by the present study (Fig. 9d; Table 1). Estimates of Lmax for C. fasciatus provided by the present study were larger than estimates of mean maximum size from visual surveys, but maximum size determined through fishery-dependent sampling and visual surveys were similar at 31.8 cm and 35 cm, respectively (Fig. 9d; Table 1). Maximum sizes for H. melapterus sampled during the present study were also similar to that observed during visual surveys in fished sites, at 40.2 cm and 45 cm, respectively (Fig. 9d; Table 1). The largest O. digramma observed in visual surveys was 24 cm in marine reserves, slightly smaller than the 26.3 cm specimen sampled during the present study, but similar to the Lmax estimated for this species (Fig. 9d; Table 1).
On the smaller end of the size-spectrum, similar minimum size classes of wrasses were purchased from fish markets during the present study to those recorded by visual census in marine reserves and fished sites (Table 1; Figs. 7, 9a–d). Furthermore, the visual census data suggest little difference in size structure of these wrasses between fished and unfished reefs in this region of the Philippines (Fig. 9d). Accordingly, fishery-dependent sampling was able to accurately represent the size structure of these wrasses in the Philippines. The size ranges of samples collected from Philippine fish markets were also similar to those collected via fishery-independent sampling on the GBR (Figs. 6, 7; Table 1; ESM 1).
Discussion
The analysis of life history variation in H. melapterus, H. fasciatus, C. fasciatus, and O. digramma sampled from Philippine fish markets and two locations on the GBR, Australia revealed several distinct trends in age structure, mortality, growth rate, and size. The present study also determined differences in the size and age of female maturity and the size and age of sex change between these three localities, and a previous study identified differences in patterns of male ontogeny of these species between the Philippines and GBR (Lowe et al. 2021b). As the studied wrasses are protogynous, these demographic traits represent additional and valuable life history features for comparison.
Differences in the life history traits of the studied wrasses manifested at different spatial scales. At biogeographic scales, between the Philippines and the GBR, extended life spans and slower initial growth in the colder region of high latitude (GBR), and earlier age at maturation in the warmer region of low latitude (Philippines) are consistent with predictions of the TSR and the expected effects of temperature on longevity. However, Philippine specimens exhibited greater relative body condition, sustained periods of growth beyond sexual maturity, and larger adult size to GBR samples. This resulted in nested growth curves which did not conform to predictions of the TSR. At regional scales, between Palm and Whitsunday reefs of the GBR, patterns of life history variation were less pronounced and differed across species, evident via the presence of both crossed and nested growth profiles. While we acknowledge that spatial correlations presented here cannot establish a causal link between environmental factors and phenotypic change, based on previous studies which investigated the effects of food resources on life history patterns (Berrigan and Charnov 1994; Huey and Kingsolver 2019), we hypothesize that increased resource levels may have had flow on effects which modified the size- and condition-related life history traits of the studied wrasses in the Philippines. Results therefore suggest that differences in food resource levels will be important in explaining the observed pattern of biogeographic life-history variation.
The postulated importance of food resources in the Philippines is derived from results of Berrigan and Charnov (1994), who demonstrated that increased food availability results in nested growth curves and rapid growth to a larger adult body size in ectotherms. Huey and Kingsolver (2019) extended this idea to investigate interactive effects of temperature and resource levels, demonstrating that ectotherms can increase their optimum operating temperature under high food availability, resulting in increased growth performance. Indeed, abundant or higher-quality food supply results in larger adult body size at low latitudes in several taxa (Conover and Present 1990; Arnett and Gotelli 2003). Certainly, in the Philippines, the observed large body size, rapid growth, and greater relative body condition suggest that benthic invertebrates, on which the focal species prey are not limiting their growth. It appears unlikely that somatic benefits are mediated by lower competition for food resources, considering that long-term densities of the studied wrasses were not consistently lower on fished Philippine reefs compared to unfished GBR reefs. Moreover, given that abundance (Russ et al. 2017) and size structure estimates for each species are similar across fished and protected sites in the central Philippines, it is unlikely that our estimates of maximum and asymptotic length were confounded by fishery-dependent sampling. Rather, given that invertebrate prey of the studied wrasses are found at higher biomass in disturbed environments (Kramer et al. 2015), Philippine reefs may be more productive than Palm and Whitsunday reefs of the GBR for two reasons; (1) the high biodiversity on Philippine reefs is thought to partly result from an intermediate level (frequency and size) of environmental disturbance (Connell 1978; Kubota and Chan 2009; McClure et al. 2021), and (2) compared to Palm and Whitsunday reefs of the GBR, which are largely uninhabited, land-use adjacent to Philippine fringing reefs likely results in higher relative nutrient runoff which is believed to benefit benthic invertebrates via a carbon flow cascade of dissolved organic material which stimulates microbial production (Cheutin et al. 2021). For these reasons, the bottom-up regulation of dietary resources may be responsible for the rapid growth rates, large adult body size, and greater relative body condition of the studied wrasses on Philippine reefs relative to those on the GBR. Ultimately, future studies are required to quantify the food supply (benthic invertebrate density) and resource intake levels for the studied wrasses on Philippine and Australian coral reefs to allow for a better understanding of the role that dietary resources play in determining life histories.
To that end, research programs that better identify and quantify species-specific resource availability, dietary composition, and dietary flexibility are required. Such programs should ideally account for spatial and temporal variation in resource availability and consumption at scales that are relevant to specific fish populations, and should expend special efforts to obtain the very small and the very large individuals in the fish population. Moreover, it is important to determine how differences in fish density may modify the expression of phenotypic differences among conspecific populations (Wood et al. 2021). These data are necessary to undertake comprehensive demographic and fitness comparisons between populations of the focal wrasses, and to determine the ecological roles of reef-fishes more generally.
Surprisingly, on Philippine reefs, the focal wrasses retained their rapid growth rate following sexual maturation, when energy is partitioned toward reproduction and away from somatic growth. This may be attained via intrinsic demographic trade-offs with reproduction, given that the studied wrasses have lower GSI’s for a given size or age in the Philippines compared to those on the GBR (Lowe et al. 2021b), or may result from extrinsic factors, such as increased resource availability or food intake on Philippine reefs. Future studies are required to more mechanistically examine the post-maturational growth patterns observed in the studied wrasses.
Demographic variation of H. melapterus, H. fasciatus, C. fasciatus, and O. digramma growth patterns, mortality rates, age of maturity, mean and maximum length and age were also detected at regional scales, between GBR reefs separated by approximately 2° of latitude. Cheilinus fasciatus and O. digramma populations on higher-latitude Whitsunday reefs grew slower, matured later, reached a larger asymptotic size, and an equal or larger maximum age than conspecifics on Palm reefs, resulting in crossed growth profiles. Conversely, H. melapterus and H. fasciatus populations on Whitsunday reefs reached lower asymptotic sizes than those on Palm reefs, exhibited reduced longevities, and H. melapterus matured earlier on Whitsunday reefs, resulting in nested growth profiles. The inconsistency of growth patterns among Palm and Whitsunday reefs for the studied wrasses suggests that factors influencing life histories at regional scales (2° latitude) may be more complex to disentangle. Nevertheless, given the hypothesized role of resources in modifying life history responses of these species between Philippine and GBR reefs, and the fact that ecological assessments showed that the focal wrasses responded to fluctuations in different benthic habitat components on Palm and Whitsunday reefs (Lowe et al. 2019, 2020), sampling of life histories across a gradient of resource variation at a regional scale will assist in elucidating the impacts of resources on life histories.
Studies which document geographic variation of reef fish demography illustrate the complex mechanisms that underlie intraspecific variation in growth rates and terminal body sizes (Trip et al. 2008; Taylor 2014; Taylor et al. 2014). On coral reefs, it is evident that numerous intrinsic (i.e., genetic, physiological) and extrinsic (i.e., environmental, ecological) factors have the potential to influence life histories across various scales. Such factors can also interact with one another and result in unexpected life history consequences, when compared to the phenotypic responses of ectotherms to the same environmental factors in isolation (see Huey and Kingsolver 2019). For this reason, it is possible that unidentified factors are interacting to result in similar life history responses of the studied wrasses between the Philippines and GBR to those expected under temperature and resource variation. For example, the truncated age structure and earlier maturation of studied wrasses in the Philippines may be a result of temperature as hypothesized, may have resulted from historical fishing pressure (Audzijonyte et al. 2016; albeit fishing did not appear to affect density or mean maximum length), may be due to density dependent processes at the social group level (Gust et al. 2002; as opposed to the similar reefal densities presented here for Philippines and GBR), or could be a result of interactive effects between additional factors for which ectotherm response norms have not yet been determined.
The problems encountered in the interpretation of the responses of ectotherms to variation in temperature are clearly illustrated by Wootton et al. (2022). Higher metabolic rates driven by increases in temperature have often been visualized as physiological limitations associated with reduced life spans, earlier sexual maturation, and reduced post-maturational growth under the TSR. Such limitations may also be reflected in temperature-associated levels of oxidative damage (Trip et al. 2016). However, the convergence of metabolic rates through acclimation in cold and warm temperature populations has focused attention on the need to consider evolutionary processes when interpreting temperature effects (Donelson et al. 2012; Wootton et al. 2022). In particular, Wootton et al., (2022) provided two major services to the interpretation of the TSR. First, the incorporation of multi-generational tests of the prevailing hypotheses illustrated the necessity of considering the adaptive significance of life-history trait variation, especially in terms of age-specific reproductive outputs, as a mechanistic driver of TSR-like life history patterns in ectotherms. However and second, moving the lens of interpretation from the laboratory to the field vastly complicates the task of identifying mechanistic processes which result in life history variation. The greater relative variation in growth rates, asymptotic length, and maximum length at local scales (e.g., among habitats) compared to large (geographical or latitudinal) scales among reef fish populations is a case in point (Choat et al. 2003; Paddack et al. 2009; Trip et al. 2008). Such results argue for the importance of adaptive responses to environmental settings strongly differentiated by disturbance and local successional features (Taylor et al. 2020). Given the context-specific nature and the likelihood of undetected interactions among factors, these issues cannot be readily separated by experimental protocols. A major point by Wootton et al. (2022) was the need for a more informed perspective on life history features of aquatic ectotherms with special reference to the level and timing of reproductive outputs mediated through development rates and age at maturation. This knowledge is critical with respect to coral reef fish populations, for which robust age-based life history estimates, including processes of reproductive ontogeny, are lacking.
At present, most hypotheses that seek to identity the drivers of demographic variation in reef fish over environmental gradients are based on correlative factors. Establishing causation will be complex and will involve additional locality-specific age-based demographic and habitat information to better identify those factors that might be subject to experimental analysis. Given the likelihood that variation in food resources may interact with temperature regimes to modify demographic responses (Huey and Kingsolver 2019), establishing standardized estimates of dietary resources for the study species is a priority. We suggest the following would provide a framework for the systematic development of such studies: (1) provide more species-specific demographic and abundance estimates of the focal species across gradients of food resources in the absence of temperature change, and across latitudinal gradients of temperature change under similar resource levels, to allow for mixed-effects modeling of demographic responses; (2) establish standardized estimates of the dietary resource levels (e.g., benthic invertebrate density) available to the studied wrasses coupled with a better understanding of resource use, dietary flexibility, and correlative effects on growth/development; (3) determine the best suite of age-based demographic variables that relate directly to organism fitness, so that demographic variation can be factored into evolutionarily important parameters.
References
Arnett AE, Gotelli NJ (2003) Bergmann’s rule in larval ant lions: testing the starvation resistance hypothesis. Ecol Entomol 28:645–650
Atkinson D (1994) Temperature and organism size: a biological law for ectotherms? Adv Ecol Res 25:1–58
Audzijonyte A, Fulton E, Haddon M, Helidoniotis F, Hobday AJ, Kuparinen A, Morrongiello J, Smith AD, Upston J, Waples RS (2016) Trends and management implications of human-influenced life-history changes in marine ectotherms. Fish Fish 17:1005–1028
Audzijonyte A, Barneche DR, Baudron AR, Belmaker J, Clark TD, Marshall CT, Morrongiello JR, van Rijn I (2019) Is oxygen limitation in warming waters a valid mechanism to explain decreased body sizes in aquatic ectotherms? Glob Ecol Biogeogr 28(2):64–77
Berrigan D, Charnov EL (1994) Reaction norms for age and size at maturity in response to temperature: a puzzle for life historians. Oikos 10(6):474–478
Brown JH, Gillooly JF, Allen AP, Savage VM, West GB (2004) Toward a metabolic theory of ecology. Ecology 85:1771–1789
Cheutin MC, Villéger S, Hicks CC, Robinson JP, Graham NA, Marconnet C, Restrepo CXO, Bettarel Y, Bouvier T, Auguet JC (2021) Microbial shift in the enteric bacteriome of coral reef fish following climate-driven regime shifts. Microorganisms 9(8):1711
Choat JH, Robertson DR, Ackerman J, Posada JM (2003) An age-based demographic analysis of the Caribbean stoplight parrotfish Sparisoma viride. Mar Ecol Prog Ser 246:265–277
Connell JH (1978) Diversity in tropical rain forests and coral reefs. Science 199:1302–1310
Conover DO, Present TM (1990) Countergradient variation in growth rate: compensation for length of the growing season among Atlantic silversides from different latitudes. Oecologia 83:316–324
Donelson JM, Munday PL, McCormick MI, Pitcher CR (2012) Rapid transgenerational acclimation of a tropical reef fish to climate change. Nat Clim Change 2(1):30–32
Gust N, Choat J, Ackerman J (2002) Demographic plasticity in tropical reef fishes. Mar Biol 140(5):1039–1051
Huey RB, Kingsolver JG (2019) Climate warming, resource availability, and the metabolic meltdown of ectotherms. Am Nat 194:E140–E150
Kramer MJ, Bellwood O, Fulton CJ, Bellwood DR (2015) Refining the invertivore: diversity and specialisation in fish predation on coral reef crustaceans. Mar Biol 162:1779–1786
Kubota H, Chan JC (2009) Interdecadal variability of tropical cyclone landfall in the Philippines from 1902 to 2005. Geophys Res Lett 36:L12802
Lek E, Fairclough DV, Hall NG, Hesp SA, Potter IC (2012) Do the maximum sizes, ages and patterns of growth of three reef-dwelling labrid species at two latitudes differ in a manner conforming to the metabolic theory of ecology? J Fish Biol 81:1936–1962
Lowe JR, Williamson DH, Ceccarelli DM, Evans RD, Russ GR (2019) Responses of coral reef wrasse assemblages to disturbance and marine reserve protection on the Great Barrier Reef. Mar Biol 166:119
Lowe JR, Williamson DH, Ceccarelli DM, Evans RD, Russ GR (2020) Environmental disturbance events drive declines in juvenile wrasse biomass on inshore coral reefs of the Great Barrier Reef. Environ Biol Fishes 103:1279–1293
Lowe JR, Payet SD, Harrison HB, Hobbs J-PA, Hoey AS, Taylor BM, Sinclair-Taylor TH, Pratchett MS (2021a) Regional versus latitudinal variation in the life-history traits and demographic rates of a reef fish, Centropyge bispinosa, in the Coral Sea and Great Barrier Reef Marine Parks, Australia. J Fish Biol 99(5):1602–1612
Lowe JR, Russ GR, Bucol AA, Abesamis RA, Choat JH (2021b) Geographic variability in the gonadal development and sexual ontogeny of Hemigymnus, Cheilinus, and Oxycheilinus wrasses among Indo-Pacific coral reefs. J Fish Biol 99(4):1348–1363
McClure EC, Hoey AS, Sievers KT, Abesamis RA, Russ GR (2021) Relative influence of environmental factors and fishing on coral reef fish assemblages. Conserv Biol 35:976–990
Munch SB, Salinas S (2009) Latitudinal variation in lifespan within species is explained by the metabolic theory of ecology. Proc Natl Acad Sci 106:13860–13864
Ogle DH, Wheeler P, Dinno A (2019) FSA: fisheries stock analysis. R package version 0.8.25.9000, https://github.com/droglenc/FSA.
Paddack MJ, Sponaugle S, Cowen RK (2009) Small-scale demographic variation in the stoplight parrotfish Sparisoma viride. J Fish Biol 75:2509–2526
Perez KO, Munch SB (2010) Extreme selection on size in the early lives of fish. Evolution 64:2450–2457
Russ GR, Alcala AC (1996) Do marine reserves export adult fish biomass? Evidence from Apo Island, central Philippines. Mar Ecol Prog Ser 132:1–9
Russ GR, Alcala AC (1998) Natural fishing experiments in marine reserves 1983–1993: community and trophic responses. Coral Reefs 17(4):383–397
Russ GR, Lowe JR, Rizzari JR, Bergseth BJ, Alcala AC (2017) Partitioning no-take marine reserve (NTMR) and benthic habitat effects on density of small and large-bodied tropical wrasses. PLoS ONE 12:e0188515
Stearns SC (1992) The evolution of life histories. Oxford University Press, Oxford
Taylor BM (2014) Drivers of protogynous sex change differ across spatial scales. Proc R Soc B Biol Sci 281:20132423
Taylor BM, Lindfield SJ, Choat JH (2014) Hierarchical and scale-dependent effects of fishing pressure and environment on the structure and size distribution of parrotfish communities. Ecography 38:520–530
Taylor BM, Benkwitt CE, Choat JH, Clements KD, Graham NA, Meekan MG (2020) Synchronous biological feedbacks in parrotfishes associated with pantropical coral bleaching. Glob Change Biol 26(3):1285–1294
Trip EL, Choat JH, Wilson DT, Robertson DR (2008) Interoceanic analysis of demographic variation in a widely distributed Indo-Pacific coral reef fish. Mar Ecol Prog Ser 373:97–109
Trip EDL, Clements KD, Raubenheimer D, Choat JH (2014) Temperature-related variation in growth rate, size, maturation and life span in a marine herbivorous fish over a latitudinal gradient. J Anim Ecol 83:866–875
Trip EDL, Clements KD, Raubenheimer D, Choat JH (2016) Temperate marine herbivorous fishes will likely do worse, not better, as waters warm up. Mar Biol 163:24
Victor BC (1986) Duration of the planktonic larval stage of one hundred species of Pacific and Atlantic wrasses (family Labridae). Mar Biol 90:317–326
Wood ZT, Lopez LK, Symons CC, Robinson RR, Palkovacs EP, Kinnison MT (2021) Drivers and cascading ecological consequences of Gambusia affinis trait variation. Am Nat. https://doi.org/10.1086/717866
Wootton HF, Morrongiello JR, Schmitt T, Audzijonyte A (2022) Smaller adult fish size in warmer water is not explained by elevated metabolism. Ecol Lett 25(5):1–12
Acknowledgements
This study was funded by the ARC Centre of Excellence for Coral Reef Studies and the College of Science and Engineering, James Cook University, Australia (JCU). We thank the Philippine Department of Agriculture—Bureau of Fisheries and Aquatic Resources (DA-BFAR), Silliman University Angelo King Centre for Research and Environmental Management (SUAKCREM), Dr. A.C. Alcala, and Emily Layos for their assistance and logistical support in the Philippines. Additional thanks to JCU Boating and Diving, Explore Group Hamilton Island, and the Orpheus Island Research Station for their operational support in Australia. A special thank you to Brock Bergseth, Gerard Ricardo, Haydn Turner, Kyle Hillcoat, Katie Sievers, Sam Smith, Roxie Diaz, and Ramon Dapat Jr. for their assistance in the field. JRL was supported by an Australian Government Research Training Program Scholarship. We sincerely thank the Associate Editor and two anonymous Reviewers for their thoughtful and constructive comments which improved this manuscript.
Funding
Open Access funding enabled and organized by CAUL and its Member Institutions. This study was funded by an ARC Centre of Excellence for Coral Reef Studies grant to GR Russ and a College of Science and Engineering, James Cook University grant to JR Lowe and GR Russ. JR Lowe was supported by an Australian Government Research Training Program Scholarship.
All applicable international, national and/or institutional guidelines for sampling, care, and experimental use of organisms for the study have been followed and all necessary approvals have been obtained. Research was carried out under James Cook University ethics permit number A2378, Great Barrier Reef Marine Park Authority collection permits G13/35909.1 and G18/38392.1, Queensland Department of Agriculture and Fisheries collection permit 170251, with approval from the Silliman University Research Ethics Committee, and in accordance with a memorandum of agreement established between JCU, Silliman University, and DA-BFAR. The authors declare that they have no conflict of interest.
Author information
Authors and Affiliations
Contributions
JR Lowe, GR Russ, JH Choat and RA Abesamis contributed to the study conception and design. Material preparation, data collection, and analysis were performed by JR Lowe, AA Bucol, RA Abesamis, and BL Stockwell. The first draft of the manuscript was written by JR Lowe and all authors commented on previous versions of the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Ethics approval
Research was carried out under James Cook University ethics permit number A2378, Great Barrier Reef Marine Park Authority collection permits G13/35909.1 and G18/38392.1, Queensland Department of Agriculture and Fisheries collection permit 170251, with approval from the Silliman University Research Ethics Committee, and in accordance with a memorandum of agreement established between JCU, Silliman University, and DA-BFAR.
Data/code availability
The majority of data underlying analysis are presented in the manuscript and supplementary materials. Additional data and code are available from the corresponding author upon reasonable request.
Additional information
Responsible Editor: S. Hamilton.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Lowe, J.R., Russ, G.R., Bucol, A.A. et al. Patterns of biogeographic and regional life-history trait variation in four large-bodied tropical wrasses. Mar Biol 169, 113 (2022). https://doi.org/10.1007/s00227-022-04098-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00227-022-04098-9